Aspergillus giganteus - Википедия - Aspergillus giganteus
| Aspergillus giganteus | |
|---|---|
| Научная классификация | |
| Королевство: | Грибы |
| Разделение: | Аскомикота |
| Учебный класс: | Евротиомицеты |
| Заказ: | Евротиалы |
| Семья: | Trichocomaceae |
| Род: | Аспергиллы |
| Разновидность: | A. giganteus |
| Биномиальное имя | |
| Aspergillus giganteus Вемер (1901)[1] | |
Aspergillus giganteus это разновидность из грибок в род Аспергиллы (от латинского aspergillum, что означает «ороситель святой водой»), который растет как плесень.[2] Впервые он был описан в 1901 году Вемером,[1] и один из шести Аспергиллы виды из Клавати раздел подрод Фумигати.[3] Ближайшие таксономические родственники: Aspergillus rhizopodus (Рай и др., 1975)[4][3] и Aspergillus longivescia (Хуанг и Рапер, 1971).[5][3]
Морфология

Как и большинство Аспергиллы разновидность,[6] A. giganteus воспроизводит бесполым путем, и не наблюдалось в сексуальном состоянии.[3] Подобен другим видам в Клавати, A. giganteus дает многочисленные конидиеносцы, которые характеризуются стеклянными и полупрозрачными стеблевыми структурами, называемыми ножками, и булавовидными аспергиллами, сужающимися к кончикам.[3] Бесполые споры сине-зеленого цвета, называемые конидии форму на этих советах.[3]
На макроскопическом уровне A. giganteus колонии характеризуются бархатистой текстурой.[3] Колонии часто сначала белые, а на свету становятся бледно-сине-зелеными.[3]

На микроскопическом уровне A. giganteus дает два типа конидиеносцев с отчетливыми ножками и пузырьками. Первые из этих конидиеносцев обычно имеют высоту 2–3 мм, включая длину ножки.[3] Эти более короткие конидиеносцы образуют булавовидные пузырьки длиной 100–250 мкм и шириной 30–50 мкм.[3] Второй тип конидиеносцев намного крупнее, примерно 1-5 см в длину.[3] Большая часть этой длины связана с удлиненными ножками.[3] Эти более удлиненные конидиеносцы фототропный, что означает, что они могут развиваться и расти только при наличии света.[3] Везикулы, продуцируемые длинными конидиеносцами, имеют длину 400-600 мкм и ширину 120-180 мкм, что более чем в два раза превышает размер везикул, производимых их более короткими аналогами.[3]
Вместе пузырек и фиалиды образуют конидиальную головку. В A. giganteus, эти головки имеют сине-зеленый цвет и разделяются на две или более колонки по мере созревания плесени.[3] Из этих конидиальных головок образуются конидии. Конидии A. giganteus имеют относительно толстые стенки и отличаются гладкой эллиптической формой, а также размером (3,5–4,5 х 2,4–3,0 мкм). Эти черты характерны для обоих типов конидиофор.[3]A. giganteus можно отличить от других Аспергиллы виды, помещенные в Клавати срез из-за его микроскопической морфологии и уникальной комбинации экстролитов, которые представляют собой соединения, синтезируемые и выделяемые клетками для защиты от бактерий и других грибов.[3][8] Морфологически, A. giganteus не хватает ризоидальных клеток стопы, присутствующих в A. rhizopodus, и имеет булавовидные пузырьки, которые контрастируют с удлиненными пузырьками A. longivesica.[3] Что касается синтеза экстролита, хотя A. giganteus производит микотоксины и антибиотики которые характерны для всех Клавати виды сечения (например, патулин,[8] триптоквивалины и триптоквивалоны,[3] и альфа-сарцины),[9] он также производит экстролиты, не синтезируемые его ближайшими родственниками. К ним относятся несколько различных каротиноиды[10] и пенициллин -подобный клавинформин.[11]
Экология
A. giganteus встречается по всему миру и официально зарегистрирован в Нигерии, США, Египте, Мексике, Панаме, Германии, Суринаме, Нидерландах и Польше.[3] Чаще всего навозный, но будет расти на щелочной почве и на деревянных субстратах.[3] Как сапротрофная плесень,[2] A. giganteus получает свою энергию, поглощая питательные вещества из субстратов, на которых растет. Хотя многие другие Аспергиллы и Клавати секционные виды - известные возбудители болезней человека, A. giganteus не имеет патогенных эффектов.[3]
В лаборатории, A. giganteus культивируется на обоих Агар с дрожжевым экстрактом Чапека (CYA) и чашки с солодовым экстрактом Agar Oxoid® (MEAOX),[3] как показано ниже.
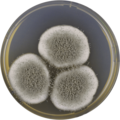
Aspergillus giganteus растет на пластине CYA
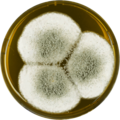
Aspergillus giganteus растет на пластине MEAOX


Экономическая актуальность
Противогрибковый белок (AFP)
A. giganteus производит противогрибковый белок (AFP), который потенциально может применяться для предотвращения грибковой инфекции как в фармацевтике, так и в сельском хозяйстве. Было показано, что AFP ограничивает рост многих экономически важных мицелиальных грибов.[12] К ним относятся Aspergillus fumigatus, основная причина аспергиллез в людях,[12] и другие Аспергиллы разновидность;[13] Fusarium oxysporum и связанные Фузариум разновидность,[12] распространенные патогены растений томата, фасоли, банана, дыни, хлопка и нута;[14] Magnaporthe grisea, возбудитель риса и зерновых культур;[2] и Botrytis cineara, патогенны для герани и других хозяйственно важных декоративных растений.[15] Кроме того, AFP может ограничивать рост оомицет Phytopthera infestans,[2] патоген, печально известный тем, что вызывает фитофтороз картофеля (основная причина Великий голод Ирландии ) и помидоры.[16]
Помимо высокого потенциала в борьбе с перечисленными выше инфекциями, AFP не подавляет жизнеспособность дрожжи, бактерии, млекопитающее,[17] или растительные клетки.[18] Поскольку существует множество видов мицелиальных грибов, которые не реагируют на АФП, пагубные эффекты белка, вероятно, зависят от вида.[13][19] Таким образом, AFP можно использовать для лечения и предотвращения инфекции очень специфическими патогенами без вреда для пациентов или растений-хозяев. Кроме того, белок можно легко синтезировать путем ферментации A. giganteus и не слышит. По сравнению с другими противогрибковыми препаратами, необходимо небольшое количество белка для предотвращения роста вредных патогенов.[20] В сочетании с тем фактом, что AFP почти полностью блокирует рост чувствительных патогенных грибов (в отличие от противогрибковых препаратов озона, перекиси водорода и диоксида хлора, используемых в настоящее время в сельскохозяйственных системах)[18] Эти факторы подчеркивают потенциал AFP как дешевого, производимого в массовом порядке и чрезвычайно эффективного решения проблемы патогенной инфекции.
Режим действия AFP
Богатый цистеином,[20] амфипатический[13] белок подавляет рост чувствительных к AFP патогенов множественными путями. Во-первых, AFP может подавлять рост клеток, подавляя хитин синтез.[12] У чувствительных грибов AFP активирует путь целостности клеточной стенки, тем самым усиливая экспрессию гена α-1,3-глюкансинтазы A, ответственного за ремоделирование клеточных стенок и подавление образования хитина.[21] АФП может также изменять проницаемость клеточной мембраны чувствительных грибов,[19] особенно Aspergillus niger,[13] или вызывают немедленное устойчивое увеличение потенциала покоя клеток с катионами кальция, что часто приводит к запрограммированной гибели клеток.[21] Каждый из этих путей в конечном итоге приводит к гибели чувствительного к АФП патогена. Несмотря на то, что необходимы дополнительные исследования, чтобы определить, как эти пути работают в консорте, большое разнообразие способов действия AFP подчеркивает, что белок может взаимодействовать с чувствительными патогенами по-разному, что эти патогены различаются по своей чувствительности к AFP.[13]
Предварительные исследования по применению AFP
В предварительных исследованиях фармацевтического и сельскохозяйственного применения AFP исследователи использовали AFP для остановки роста Phytopthera infestans[2] и Аспергиллы разновидность[12][13][19] в культуре. При нанесении растворов AFP на листья и корни растений инфекции Botrytis cineara в растениях герани[15] и Magnaporthe grisea в растениях риса были устранены.[2] В каждом из этих случаев AFP вызывал деформации гиф чувствительных грибов, что приводило к уменьшению удлинения гиф и неспособности патогена к росту.[2][15] В дополнение к прямому нанесению на структуры растений ген, кодирующий AFP, можно редактировать в геномах растений, пораженных AFP-чувствительными грибами, так что растения могут синтезировать белок самостоятельно.[22][23][24] Этот метод был успешно использован для уменьшения заражения Puccinia substriata, основная причина болезни ржавчины, и Sclerospora graminicola, возбудитель ложной мучнистой росы в растениях перлового проса,[22] а также заражение Magnaporthe grisea в рисовых растениях.[23][24] Хотя генетическое улучшение остановило инфекцию в лабораториях и теплицах, исследователи продолжают спорить о масштабируемости редактирования генома растений и способности самих растений синтезировать достаточное количество AFP для борьбы с инфекцией патогенов в полевых условиях.[23][24]
Рекомендации
- ^ а б Wehmer 1901, Mem. Soc. Phys. Женева 33 (2): 85
- ^ а б c d е ж грамм Vila L, Lacadena V, Fontanet P, Martinez del Pozo A, San Segundo B (ноябрь 2001 г.). «Белок плесени Aspergillus giganteus - мощный ингибитор грибковых патогенов растений». Молекулярные взаимодействия растений и микробов. 14 (11): 1327–31. Дои:10.1094 / MPMI.2001.14.11.1327. PMID 11763131.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс Варга Дж., Дуэ М., Фрисвад Дж. К., Самсон РА (2007). «Таксономическая ревизия Aspergillus секции Clavati на основе молекулярных, морфологических и физиологических данных». Исследования в области микологии. 59: 89–106. Дои:10.3114 / sim.2007.59.11. ЧВК 2275193. PMID 18490946.
- ^ Рай Дж. Н., Вадхвани К., Агарвал СК (июнь 1975 г.). «Aspergillus rhizopodus sp.nov. Из индийских солонцеватых почв». Труды Британского микологического общества. 64 (3): 515–517. Дои:10.1016 / с0007-1536 (75) 80153-7. ISSN 0007-1536.
- ^ Хуан Л.Х., Рэпер КБ (январь 1971 г.). «Aspergillus Longivesica, новый вид из нигерийской почвы». Микология. 63 (1): 50–57. Дои:10.1080/00275514.1971.12019081. ISSN 0027-5514.
- ^ Дайер П.С., Паолетти М. (май 2005 г.). "Размножение Aspergillus fumigatus: сексуальность предположительно бесполого вида?". Медицинская микология. 43 Дополнение 1 (s1): S7-14. Дои:10.1080/13693780400029015. PMID 16110786.
- ^ «Аспергиллы». Микология онлайн. Университет Аделаиды. Получено 2019-05-17.
- ^ а б Варга Дж., Риго К., Мольнар Дж., Тот Б., Сенч С., Терен Дж., Козакевич З. (2003). «Производство микотоксинов и эволюционные отношения между видами Aspergillus секции Clavati». Антони ван Левенгук. 83 (2): 191–200. Дои:10.1023 / А: 1023355707646. PMID 12785313.
- ^ Линь А., Хуан К.С., Хву Л., Цзян СС (январь 1995 г.). «Производство риботоксинов типа II видами Aspergillus и родственными грибами на Тайване». Токсикон. 33 (1): 105–10. Дои:10.1016/0041-0101(94)00140-4. PMID 7778122.
- ^ ван Эйк Г.В., Маммери Р.С., Ройманс Г.Дж., Валадон Л.Р. (1979-09-01). «Сравнительное исследование каротиноидов Aschersonia aleyroides и Aspergillus giganteus». Антони ван Левенгук. 45 (3): 417–422. Дои:10.1007 / BF00443280. ISSN 1572-9699. PMID 554534.
- ^ Флори HW, Дженнингс MA, Philpot FJ (январь 1944 г.). «Клавиформин из Aspergillus giganteus Wehm». Природа. 153 (3874): 139. Bibcode:1944Натура.153..139F. Дои:10.1038 / 153139a0.
- ^ а б c d е Хаген С., Маркс Ф, Рам А.Ф., Мейер В. (апрель 2007 г.). «Противогрибковый белок AFP из Aspergillus giganteus подавляет синтез хитина у чувствительных грибов». Прикладная и экологическая микробиология. 73 (7): 2128–34. Дои:10.1128 / AEM.02497-06. ЧВК 1855660. PMID 17277210.
- ^ а б c d е ж Мейер V (февраль 2008 г.). «Небольшой белок, который борется с грибками: AFP как новый многообещающий противогрибковый агент, имеющий биотехнологическую ценность». Прикладная микробиология и биотехнология. 78 (1): 17–28. Дои:10.1007 / s00253-007-1291-3. PMID 18066545.
- ^ Michielse CB, Rep M (май 2009 г.). «Обновление профиля патогена: Fusarium oxysporum». Молекулярная патология растений. 10 (3): 311–24. Дои:10.1111 / j.1364-3703.2009.00538.x. ЧВК 6640313. PMID 19400835.
- ^ а б c Морено А.Б., Дель Посо А.М., Борха М., Сегундо Б.С. (ноябрь 2003 г.). «Активность противогрибкового протеина из Aspergillus giganteus против Botrytis cinerea». Фитопатология. 93 (11): 1344–53. Дои:10.1094 / PHYTO.2003.93.11.1344. PMID 18944061.
- ^ Баркай-Голан Р (июнь 2001 г.). Послеуборочные болезни фруктов и овощей (1-е изд.). Эльзевир. ISBN 978-0-444-50584-2.
- ^ Szappanos H, Szigeti GP, Pál B, Rusznák Z, Szucs G, Rajnavölgyi E, Balla J, Balla G, Nagy E, Leiter E, Pócsi I, Hagen S, Meyer V, Csernoch L (июль 2006 г.). «Противогрибковый белок AFP, секретируемый Aspergillus giganteus, не оказывает вредного воздействия на определенные клетки млекопитающих». Пептиды. 27 (7): 1717–25. Дои:10.1016 / j.peptides.2006.01.009. PMID 16500727.
- ^ а б Баракат Х., Шпильвогель А, Хассан М, Эль-Десуки А, Эль-Манси Х., Рат Ф., Мейер В., Шталь У. (июнь 2010 г.). «Противогрибковый белок AFP из Aspergillus giganteus предотвращает вторичный рост различных видов Fusarium на ячмене». Прикладная микробиология и биотехнология. 87 (2): 617–24. Дои:10.1007 / s00253-010-2508-4. PMID 20217075.
- ^ а б c Theis T., Wedde M, Meyer V, Stahl U (февраль 2003 г.). «Противогрибковый белок из Aspergillus giganteus вызывает мембранную проницаемость». Противомикробные препараты и химиотерапия. 47 (2): 588–93. Дои:10.1128 / aac.47.2.588-593.2003. ЧВК 151754. PMID 12543664.
- ^ а б Лакадена Дж., Дель Посо А.М., Гассет М., Патино Б., Кампос-Оливас Р., Васкес С. и др. (Декабрь 1995 г.). «Характеристика противогрибкового белка, секретируемого MouldAspergillus giganteus». Архивы биохимии и биофизики. 324 (2): 273–81. Дои:10.1006 / abbi.1995.0040. PMID 8554319.
- ^ а б Биндер У., Бенсина М., Эйгентлер А., Мейер В., Маркс Ф. (сентябрь 2011 г.). «Противогрибковый белок Aspergillus giganteus AFPNN5353 активирует путь целостности клеточной стенки и нарушает гомеостаз кальция». BMC Microbiology. 11 (1): 209. Дои:10.1186/1471-2180-11-209. ЧВК 3197501. PMID 21943024.
- ^ а б Гирджи М., Бриз В.А., Лёрц Х., Олдах К.Х. (июнь 2006 г.). «Устойчивость к ржавчине и ложной мучнистой росе жемчужного проса (Pennisetum glaucum), опосредованная гетерологичной экспрессией гена afp из Aspergillus giganteus». Трансгенные исследования. 15 (3): 313–24. Дои:10.1007 / s11248-006-0001-8. PMID 16779647.
- ^ а б c Кока М., Бортолотти С., Руфат М., Пеньас Дж., Эритха Р., Тарро Д., дель Посо А.М., Мессегер Дж., Сан-Сегундо Б. (январь 2004 г.). «Трансгенные растения риса, экспрессирующие противогрибковый белок AFP из Aspergillus giganteus, демонстрируют повышенную устойчивость к рисовому грибку Magnaporthe grisea». Молекулярная биология растений. 54 (2): 245–59. Дои:10.1023 / B: PLAN.0000028791.34706.80. PMID 15159626.
- ^ а б c Морено А.Б., Пеньас Дж., Руфат М., Браво Дж. М., Эстопа М., Мессегер Дж., Сан-Сегундо Б. (сентябрь 2005 г.). «Вызванное патогенами производство противогрибкового белка AFP из Aspergillus giganteus придает устойчивость к бластному грибку Magnaporthe grisea в трансгенном рисе». Молекулярные взаимодействия растений и микробов. 18 (9): 960–72. Дои:10.1094 / MPMI-18-0960. PMID 16167766.