Хронология исследований гадрозавров - Википедия - Timeline of hadrosaur research

Этот хронология исследований гадрозавров хронологический список событий в история из палеонтология сосредоточился на гадрозавроиды, группа травоядный орнитопод динозавры широко известный как утконосые динозавры. Научные исследования гадрозавров начались в 1850-е годы,[1] когда Джозеф Лейди описал роды Теспезий и Trachodon на основе лоскутных окаменелостей обнаруженный в западный Соединенные Штаты. Всего два года спустя он опубликовал описание гораздо лучше сохранившихся останков животного. из Нью-Джерси что он назвал Гадрозавр.[2]
Рано 20 век увидел такой бум открытий и исследований гадрозавров, что знания палеонтологов об этих динозаврах «увеличились практически на порядок», согласно 2004 обзор Хорнер, Weishampel, и Форстер. Этот период известен как великая атака динозавров в Северной Америке из-за исследований и раскопок таких палеонтологов, как коричневый, Гилмор, Ламбе, Парки, а Штернбергс. Основные открытия включали в себя разнообразие черепных украшений у гадрозавров, поскольку ученые стали охарактеризовать некристальные, твердые и полые гребешковые виды.[2] Включены заметные новые таксоны Зауролоф, Коритозавр, Эдмонтозавр, и Ламбеозавр.[3] В 1942 Ричард Суонн Затишье и Райт опубликовали то, что Хорнер, Вейшампель и Форстер охарактеризовали как «первый важный синтез анатомии и филогении гадрозаврид».[2]
Более поздние открытия включают гигантских гадрозавров, таких как Шантунгозавр гигантский из Китай.[4] При длине 15 метров и весе около 16 метрических тонн это самый крупный из известных гадрозавров, известный по почти полному скелету.[5]
Исследования хадрозавров продолжаются даже в новом тысячелетии. В 2000, Хорнер и другие обнаружили, что детеныш Майасаура вырос до размеров взрослого тела со скоростью, больше похожей на млекопитающее чем рептилия. В том же году Кейс и другие сообщили об обнаружении костей гадрозавра в Остров Вега, Антарктида. После десятилетий таких целенаправленных исследований гадрозавры стали одной из наиболее изученных групп динозавров.[2]
19 век
1850-е годы



- Джозеф Лейди описал новый род и вид Thespesius occidentalis. Он также описал новый род и вид Trachodon mirabilis.[1] Хотя оба вида были основаны на плохо сохранившемся материале, эта статья была первой опубликованной по динозаврам гадрозаврид.
- Лейди описал новый род и вид Hadrosaurus foulkii.[6] Он думал, что это животное-амфибия.[7]
1860-е годы
- Лейди сотрудничал с художником Бенджамин Уотерхаус Хокинс монтировать Hadrosaurus foulkii для Академия естественных наук Филадельфии. Это был первый скелет динозавра, когда-либо выставленный для всеобщего обозрения, а также один из самых популярных экспонатов в истории Академии. Оценки имеют Гадрозавр выставка как увеличение количества посетителей до 50%.[8]
- Эдвард Дринкер Коуп описал новый род и вид Hypsibema crassicauda.[9]
- Коуп назвал Hadrosauridae. Он заметил, что основной отличительной чертой группы была их стоматологическая батарея.[10]
- Коп описал новый род и вид Орнитотарсус имманис.[6]
1870-е годы
- Отниэль Чарльз Марш описал новый вид Гадрозавр малый.
- Коп описал новый вид Hadrosaurus cavatus.
- Марш описал новый вид Hadrosaurus agilis.[11]
- Коп описал новый вид Агатаумас Мило.[6] Он также описал новый род и вид Ционодон арктатус.[9]
- Коп описал новый вид Cionodon stenopsis.[9]
- Коп описал новый род и вид Диклоний кальмар. Он также описал новый вид Диклоний пятиугольный, Диклониус перангулатус, и Дисган энкаустус.[9]
1880-е годы


- Гарри Говьер Сили описал новый род и вид Orthomerus dolloi.[1]
- Коп все еще считал гадрозавров амфибиями.[7]
- Ричард Лидеккер описал новый вид Trachodon cantabrigiensis.[1]
- Марш описал новый вид Hadrosaurus breviceps. Он также описал новый вид Hadrosaurus paucidens.[9]
- Коп описал новый род и вид Птеропеликсовые гралипсы.[1]
1890-е годы
- Марш возвел новый род Клаозавр разместить вид Hadrosaurus agilis.[11] Он также описал новый вид Траходон длинноглавый.[6]
- Марш описал новый вид Claosaurus annectens.[6]
- Коп описал новый род и вид Claorhynchus trihedrus.[9]
- Ньютон описал новый вид Игуанодон холмистый.[9]
20 век
1900-е годы
- Франц Нопца описал новый род и вид Лимнозавр транссильванский.[11]
- Лоуренс Ламбе описал новый род и вид Trachodon altidens; в том же томе он предложил название Диданодон для того же вида, но правомерность этого названия была поставлена под сомнение.[12][9] Он также описал новый вид Trachodon marginatus. Он также описал новый вид Trachodon selwyni.[1]
- Nopcsa возвел новый род Телматозавр разместить вид Лимнозавр транссильванский, так как последнее название рода было озабочено.[11]
- Джордж Виланд описал новый вид Claosaurus affinis.[9]
1910


- Барнум Браун описал новый род Гекатазавр.[11] Он также описал новый род и вид Kritosaurus navajovius.[9]
- Браун описал новый род и вид Saurolophus osborni.[4]
- Браун описал новый род и вид Гипакрозавр альтиспинус.[4]
- Катлер раскопал несовершеннолетнего Грипозавр теперь внесен в каталог Канадского музея природы как CMN 8784. Место раскопок с тех пор обозначено как «карьер 252».[13]
- Зима: Катлер частично подготовил молодых Грипозавр Образец, возможно, в Калгари, когда работал над динозаврами для Юстона Сисели.[13]
- Браун описал новый род и вид Коритозавр казуарий.[4]
- Ламбе описал новый род и вид Gryposaurus notabilis.[6]
- Чарльз Х. Штернберг бригада раскопала Коритозавр из карьера 243 в Провинциальном парке динозавров, Альберта, Канада. Образец позже будет выставлен в зоопарке Калгари.[14]
- Мэтью заметил, что окаменелости гадрозавра яйца птенцы отсутствовали в прибрежных районах, что позволяет предположить, что гадрозавры могли предпочитать места гнездования дальше вглубь суши. Он считал, что эти внутренние районы гнездования на самом деле были местом, где гадрозавры впервые эволюционировали, и поэтому для размножения гадрозавры проследили путь своих предков до места своего происхождения. После вылупления молодые гадрозавры проводят некоторое время в глубине суши, созревая, прежде чем мигрировать в более прибрежные районы.[15]

- Браун описал новый род и вид Prosaurolophus maximus.[6]
- Ламбе описал новый род и вид Эдмонтозавр королевский.[6]
- Ламбе описал новый род и вид Cheneosaurus tolmanensis.[4]
- Ламбе назвал Hadrosaurinae.
1920-е годы


- Мэтью описал новый род Procheneosaurus.[9]
- Парки описали новый вид Критозавр incurvimanus.[6]
- Уильям Паркс описал новый род и вид Parasaurolophus walkeri.[9]
- Краузель сообщил об ископаемом содержимом кишечника Эдмонтозавр аннектенс Мама. Он описал материал как включающий хвойное дерево иголки и ветки, лиственный листва, а возможно и мелкая семена или же фрукты.[7]
- Абель утверждал, что растительный материал, который, как утверждает Краузель, был окаменелыми остатками содержимого кишечника Эдмонтозавр аннектенс был фактически нанесен проточной водой.[7]
- Чарльз Уитни Гилмор описал новый вид Коритозавр земной.[4]
- Парки описали новый вид Коритозавр промежуточный.[4] Он также описал новый род и вид Ламбеозавр ламбей.[9]
- Гилмор описал новый вид Thespesius edmontonensis.[6]

- Рябинин описал новый вид Trachodon amurensis.[1]
- Штернберг описал новый вид Thespesius saskatchewanensis.[6]
- Виман описал новый род и вид Tanius sinensis.[11]
1930-е годы

- Рябинин описал новый вид Saurolophus kryschtofovici.[1]
- Рябинин возвел новый род Мандшурозавр разместить вид Trachodon amurensis.[1]
- Паркс описал новый род и вид Тетрагонозавр прямостоячий.[4] Он также описал новый вид Прецепс тетрагонозавра.
- Виман описал новый вид Parasaurolophus tubicen.[9]
- Рябинин описал новый род и вид. Ционодон kysylkumensis.[9]
- Гилмор описал новый род и вид Бактрозавр johnsoni.[11] Он также описал новый вид Mandschurosaurus mongoliensis.[11]
- Парки описали новый вид Коритозавр bicristatus и C. brevicristatus.[4]

- Штернберг описал новый вид Тетрагонозавр кранибревис.[4] Он также описал новый вид Ламбеозавр clavinitialis и L. magnicristatus.[9]
- Парки описали новый вид Коритозавр фронтальный.
- Такуми Нагао описал новый род и вид Nipponosaurus sachalinensis.[9]
- Рябинин описал новый род и вид. Jaxartosaurus aralensis.[4] Он также описал новый вид Бактрозавр prynadai.[9]
1940-е годы

- Ричард Суонн Затишье и Райт описал новый род Анатозавр за Claosaurus annectens. Они также называют новый вид Анатозавр копейский.[6]
- Хоффет описал новый вид Мандшурозавр лаосенсис.[1]
- Рябинин описал новый вид Orthomerus weberi.[1]
- Гилмор и Стюарт описали новый род и вид Неозавр миссурианский.[1]
- Гилмор возвел новый род Паррозавр разместить вид Неозавр миссурианский, как имя Неозавр был занят.[1]
- Янг описал новый род и вид Санпасавр яой.[1]
- Кольбер переопределил вид Гадрозавр малый.[4]
1950-е годы

- Рождественский описал новый вид Saurolophus angustirostris.[4]
- Штернберг описал новый род и вид Брахилофозавр канадский.[11]
- Янг описал новый вид Таниус чингканкоуэнсис.[11]
- Янг описал новый род и вид Цинтаозавр spinorhinus.[9]
1960-е

- Лэнгстон описал новый род и вид Лофорхотон атопус.[6]
- Остром описал новый вид Parasaurolophus cyrtocristatus.[9]
- Остром поддержал заявление Краузеля 1922 года о том, что ископаемый растительный материал, связанный с Эдмонтозавр аннектенс мумие на самом деле было его внутренним содержимым.[7]
- Рассел и Чамни изучали распространение гадрозавров в маастрихтской Северной Америке. Был сделан вывод, что Эдмонтозавр королевский жил у берегов, пока Гипакрозавр альтиспинус и Saurolophus osborni жили немного дальше от суши.[15]
- Рождественский описал новый род и вид Аралозавр трубчатый.[11] Он также описал новый вид Procheneosaurus убеждает.[4]
1970-е годы
- Гальтон утверждал, что анатомия таза гадрозавра больше соответствовала горизонтальной позе, подобной той, что наблюдается у современных нелетающих птиц, чем позе «кенгуру», в которой они часто реконструируются.[7]
- Додсон утверждал, что гадрозавры, возможно, не питались исключительно землей.[7]
- Ху описал новый род и вид Шантунгозавр гигантский.[4]

- Додсон с помощью морфометрии обнаружил доказательства полового и онтогенетического диморфизма у двух разных видов ламбеозаврина.[16]
- Чжэнь описал новый вид Таниус Лайенгенсис.[9]
- Бретт-Сурман создал новый род Гилмореозавр разместить вид Mandschurosaurus mongoliensis. Он также описал новый род и вид Secernosaurus koerneri.[11]
- Бретт-Сурман не смог определить, где впервые появились гадрозавры.[15]
- Хорнер и Макела описали новый род и вид Maiasaura peeblesorum.[6] Они утверждали, что гадрозавры заботились о своих детенышах в течение длительного периода после вылупления.[16]
- Хорнер утверждал, что окаменелости гадрозавров, найденные в морских отложениях, были просто сохранившимися останками людей, которые были смыты в море из наземного места происхождения.[7]
- Донг описал новый род и вид Microhadrosaurus nanshiungensis.[1]
1980-е

- Хоттон утверждал, что некоторые гадрозавры, возможно, мигрировали сезонно в направлении с севера на юг.
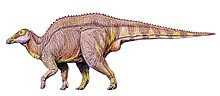

- Тереза Марьянская и Осмольская описали новый род и вид Барсболдия сицинская.[4]
- Моррис описал новый вид Lambeosaurus laticaudus.[9]
- Суслов и Шилин описали новый род и вид. Arstanosaurus akkurganensis.[9]
- Карпентер оспаривал идею о том, что гадрозавры гнездятся только на возвышенностях, вместо этого утверждая, что ископаемые яйца и детеныши гадрозавров отсутствовали только в прибрежных отложениях, потому что химический состав древних почв был просто слишком кислым, чтобы их сохранить.[7]
- Талборн утверждал, что гадрозавры могли бегать со скоростью до 14–20 км / ч в течение продолжительных периодов времени.[7]
- Хорнер заметил, что Maiasaura peeblesorum известно только, что они жили в верхних частях современных прибрежных равнин.[15]
- Вейшампель описал жевательную и черепную кинетику гадрозавров.[7]
- Weishampel и Weishampel[требуется разъяснение ] сообщили о наличии останков гадрозавров на Антарктическом полуострове.[15]
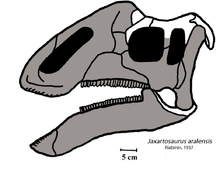

- Ву описал новый род и вид Джаксартозавр fuyuensis.[9]
- Милнер и Норман утверждали, что гадрозавры произошли в Азии.[15]
- Хорнер заметил, что окаменелые яйца и вылупившиеся детеныши гадрозавров были обычным явлением в отложениях, отложившихся в верхних областях того, что когда-то было прибрежными равнинами.[15]
- Вейшампель описал жевательную и черепную кинетику гадрозавров.[7]
- Норман описал жевательную и черепную кинетику гадрозавра.[7]
- Вайсхампель утверждал, что гадрозавры питались в основном растительностью 2 м высотой или меньше, но имели максимальную высоту прохождения 4 м.[7]
- Бонапарт и другие описали новый вид Kritosaurus australis.[4]
- Норман и Вейшампель описали гадрозавр жевательную и черепную кинетику.[7]
- Хорнер заметил, что окаменелые яйца и вылупившиеся детеныши гадрозавров были обычным явлением в отложениях, отложившихся в верхних областях того, что когда-то было прибрежными равнинами.[15]
- Фарлоу утверждал, что их высокоразвитые жевательные способности и большой объем кишечника означают, что гадрозавры были в высшей степени приспособлены к питанию бедной питательными веществами волокнистой растительностью.[7]
- Хорнер описал новый вид Брахилофозавр goodwini.[11]
1990-е
- Бретт-Сурман описал новый род Анатотитан за Анатозавр копейский.
- Хорнер утверждал, что гадрозавриды не являются естественной группой, а вместо этого две основные группы гадрозавров, обычно неребристые гадрозаврины и гребенчатые ламбеозавры имеют разные корни в пределах Игуанодонтия. Хорнер считал, что некрещеные гадрозавры произошли от родственника Игуанодон, а хохлатые ламбеозавры произошли от родственника Уранозавр. Однако это предложение не нашло поддержки ни в одной последующей исследовательской публикации.[10]
- Вейшампель и Хорнер обнаружили, что Hadrosauridae в конце концов являются естественной группой.[10] Они также нашли кладистическую поддержку традиционного разделения Hadrosauridae на подсемейства Hadrosaurinae и Lambeosaurinae.[10]
- Вайшампель сообщил о присутствии гадрозавров на Антарктическом полуострове.[15][требуется разъяснение ]
- Болоцкий и Курзанов описали новый род и вид Амурозавр риабинини.[4]

- Хорнер описал новый вид Грипозавр латиденс. Он также описал новый вид Prosaurolophus blackfeetensis.[6]
- Хант и Лукас описали новый род и вид Анасазизавр горнери.[11] Они также описали новый род и вид Naashoibitosaurus ostromi.[6]
- Вайшампель, Норман и Гриогеску назвали кладу Эухадрозаврия.[11]
- Weishampel и другие предложили определение Hadrosauridae на основе узлов: потомки последнего общего предка, разделяемые Телматозавр и Паразауролоф.[17] Они обнаружили, что гадрозавры являются естественной группой, вопреки аргументам Хорнера 1990 года о том, что подсемейства гадрозавров произошли от разных видов игуанодонтов.[10] Они также нашли кладистическую поддержку традиционного разделения Hadrosauridae на подсемейства Hadrosaurinae и Lambeosaurinae.[10]
- Клаус и Хорнер сообщили о наличии окаменелостей, зародышей и вылупившихся гадрозавров. Формация реки Джудит Монтаны. Поскольку эти отложения отложились на низменной прибрежной равнине, открытие исследователей противоречило предыдущим гипотезам о том, что гадрозавры либо не гнездились в низинах, либо местная древняя почва была слишком кислой, чтобы их сохранить.[7]

- Хорнер и Карри описали новый вид Гипакрозавр stebingeri.[4]
- Чин и Гилл описали Maiasaura peeblesorum копролиты из древнего гнездовья этого вида. Копролиты представляли собой «глыбовые» массы неправильной формы, сохранившие фрагменты растений. Исследователи определили это как фекалии, потому что массы содержали окаменелые норы навозных жуков. Растительный материал предполагал диету, состоящую в основном из стеблей хвойных деревьев.[7]
- Форстер обнаружил, что гадрозавры являются естественной группой, вопреки аргументам Хорнера 1990 года о том, что подсемейства гадрозавров произошли от разных видов игуанодонтов.[10] Они также нашли кладистическую поддержку традиционного разделения Hadrosauridae на подсемейства Hadrosaurinae и Lambeosaurinae.[10] Она предпочла определить Hadrosauridae как самых недавних общих предков гадрозавринов и ламбеозавринов и всех их потомков. В отличие от определения, использованного Вейшампелем и другими в 1993 г., это определение исключало Телматозавр.[18]
- Серено обнаружил, что гадрозавры являются естественной группой, вопреки аргументам Хорнера 1990 года о том, что подсемейства гадрозавров произошли от разных видов игуанодонтов.[10]
21-го века
2000-е
- Годфройт, Зан и Джин описали новый род и вид Charonosaurus jiayinensis.[4]
- Кейс и другие сообщили о присутствии гадрозавров на полуострове Антарктида.[2] Исследованные останки были найдены на Остров Вега и представляют собой самые южные из известных окаменелостей гадрозавров. Когда животные были еще живы, это место, вероятно, находилось примерно на 65 градусах южной широты.[15]
- Хорнер и другие исследовали гистологию Maiasaura peeblesorum кости. Они обнаружили, что Майасаура Чтобы достичь размера взрослого человека, потребовалось всего 8–10 лет. Взрослый 7 метров (23 фута) Майасаура может иметь массу тела взрослого человека более 2000 килограммов (4400 фунтов), несмотря на то, что вылупился при длине около полуметра и массе тела менее килограмма. Это несоответствие подразумевает скорость или рост, аналогичный тем, которые наблюдаются у современных млекопитающих.[7]

- Хорнер и другие опубликовали дополнительные исследования по гистологии Maiasaura peeblesorum кости.[7]
- Вы и другие описали новый род и вид Equijubus normani.[19]
- Кобаяши и Адзума описали новый род и вид Fukuisaurus tetoriensis.[20]
- Годфройт, Болотский и Алифанов описали новый род и вид. Олоротитан архаренсис.[9]

- Болотский и Годфройт описали новый род и вид Керберозавр манакини.[21]
- Годфройт, Ли и Шан описали новый род и вид. Penelopognathus weishampeli.[22]
- Прието-Маркес и другие описали новый род и вид Кутализавр kohlerorum.[23]
- Гилпин и другие описали новый род и вид Cedrorestes crichtoni.[24]

- Мо и другие описали новый род и вид Наннингозавр dashiensis.[25]
- Чжао и другие описали новый род и вид Zhuchengosaurus maximus.[26]
- Годфройт и другие описали новый род и вид Sahaliyania elunchunorum и новый род и вид Wulagasaurus dongi.[27]
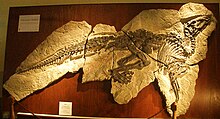
- Вагнер и Леман описали новый род и вид Ангуломастакатор daviesi.[28]
- Переда-Субербиола и др. Описали новый род и вид. Аренизавр ардеволи.[29]
- Сьюс и Аверьянов описали новый род и вид. Levnesovia transoxiana.[30]
- Далла Веккья описала новый род и вид Tethyshadros insularis.[31]
2010-е
- Крузадо-Кабальеро и другие описали новый род и вид Blasisaurus canudoi.[32]
- Прието-Маркес описал новый род и вид Glishades ericksoni.[33]
- Хуарес Вальери и другие описали новый род и вид Willinakaqe salitralensis.[34]
- Гейтс и другие описали новый род и вид Acristavus gagslarsoni.[35]
- Годфройт и другие описали новый род и вид Батырозавр Рождественский.[36]
- Рамирес-Веласко и другие описали новый род и вид Huehuecanauhtlus tiquichensis.[37]
- Годфройт и другие описали новый род и вид Кундурозавр нагорный.[38]
- Кориа, Рига и Касадио описали новый род и вид Lapampasaurus cholinoi.[39]
- Прието-Маркес и Браньяс описали новый род и вид Latirhinus uitstlani.[40]
- Прието-Маркес Чиаппе и Джоши описали новый род и вид Magnapaulia.[41]
- Прието-Маркес и другие описали новый род и вид Canardia garonnensis.[42]
- Фил Р. Белл и Кирстин С. Бринк описали новый род и вид. Kazaklambia убеждает.[43]
- Прието-Маркес и Вагнер описали новый вид Saurolophus morrisi.[44]
- Ван и другие описали новый род и вид Юнганлонг datongensis.[45]
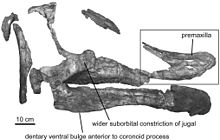
- Прието-Маркес и другие описали новый род Августинолоф.[46]
- Гейтс и Шитц описали новый род и вид Кондруп Ринорекса.[47]
- Син и другие описали новый род и вид Zhanghenglong yangchengensis.[48]
- Гейтс и другие описали новый род Адельолоф.[49]
- Вы, Ли и Додсон описали новый род Гонгпокванзавр.[50]

- Шибата и Адзума описали новый род и вид Кошизавр кацуяма.[51]
- Мори, Дракенмиллер и Эриксон описали новый род и вид Угрунаалук куукпикенсис.[52]
- Фридман Фаулер и Хорнер описали новый род и вид. Пробрахилофозавр.[53]
- Шибата и другие описали новый род и вид Сириндхорна хоратенсис.[54]
- Сюй и другие описали новый род и вид Датунлун.[55]
- Ван и другие описали новый род и вид Цзоюньлун.[56]
- Прието-Маркес, Эриксон и Эберсол описали новый род и вид Eotrachodon orientalis[57]
- Крузадо-Кабальеро и Пауэлл описали новый род и вид Bonapartesaurus rionegrensis.
- Изучение корполитов Чином, Фельдманном и Ташманом показывает, что гадрозавры иногда потребляли гниющую древесину и ракообразных
- Гейтс и другие описали новый род и вид Чойродон барсболди.[58]
- Прието-Маркес и другие описали новый род и вид Адиномозавр арканус.[59]
- Чжан и другие описали новый род и вид Лайянгозавр молодой.[60]
- Цогтбаатар и другие описали новый род и вид Gobihadros mongoliensis.[61]
- Прието-Маркес, Вагнер и Леман описали новый род и вид Aquilarhinus palimentus.[62]
- Кобаяши и другие описали новый род и вид Kamuysaurus japonicus.
- Исследование природы речной системы Ларамидия в позднем меловом периоде, о чем свидетельствуют данные по окаменелостям позвоночных и беспозвоночных из Кайпаровицкая свита южных Юта, и от поведения гадрозаврид динозавров над этими ландшафтами, будет опубликовано Crystal и другие. (2019).[63]
- Исследование по остеология и филогенетические отношения "Таниус laiyangensis " публикует Чжан и другие. (2019).[64]
- Этюд на кости гистология из большеберцовые кости из Майасаура Peeblesorumс акцентом на составе, частоте и корковой степени локализованных сосудистых изменений, опубликовано Woodward (2019).[65]
- Три молодых особи Прозавролоф Максимус, предоставляя новую информацию о онтогенез этого таксона, описаны с Формация Медвежьей Лапы (Альберта, Канада ) от Драйсдейл и другие. (2019).[66]
- Исследование влияния структуры костной ткани на раннем этапе диагенетический режимы и другие тафономический переменные по потенциалу сохранения мягких тканей в окаменелостях позвоночных, о чем свидетельствуют данные окаменелостей Эдмонтозавр аннектенс со стоянки хадрозавров в Стоячей скале (Маастрихтский Формация Адского ручья, южная Дакота ), опубликовано Ullmann, Pandya & Nellermoe (2019), которые сообщают о первом восстановлении остеоциты и сосуды из окаменелости позвоночный центр и окостеневший сухожилия.[67]
- Первый окончательный ламбеозаврин ископаемое (изолированная кость черепа) описывается из костного ложа Лискомб Формация Принс Крик (Аляска, Соединенные Штаты ) от Такасаки и другие. (2019).[68]
- Сохранились следы на хвостовом позвонке гадрозаврид динозавр из Верхний мел Формация Адского ручья (Монтана, Соединенные Штаты ) описаны Peterson & Daus (2019), которые интерпретируют свою находку как следы кормления, произведенные молодью поздней стадии тиранозавр Рекс.[69]
Смотрите также
Сноски
- ^ а б c d е ж грамм час я j k л м п о Хорнер, Вайшампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», стр. 443.
- ^ а б c d е Хорнер, Вайсхампель и Форстер (2004); «Введение», стр. 438.
- ^ Хорнер, Вайшампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», страницы 439–442.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Хорнер, Вайшампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», стр. 441.
- ^ Лукас (2001); «Немегтийские позвоночные», стр. 181.
- ^ а б c d е ж грамм час я j k л м п о п Хорнер, Вайшампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», стр. 440.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Хорнер, Вайсхампель и Форстер (2004); «Палеоэкология, биогеография и палеобиология», стр. 462.
- ^ Вайшампель и Янг (1996); "Хэддонфилд Гадрозавр", стр. 71.
- ^ а б c d е ж грамм час я j k л м п о п q р s т ты v ш Икс у Хорнер, Вайсхампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», стр. 442.
- ^ а б c d е ж грамм час я Хорнер, Вайшампель и Форстер (2004); «Систематика и эволюция», стр. 457.
- ^ а б c d е ж грамм час я j k л м п о Хорнер, Вайсхампель и Форстер (2004); «Таблица 20.1: Hadrosauridae», стр. 439.
- ^ Лунд, Э. и Гейтс, Т. (2006). «Историческое и биогеографическое исследование гадрозавров динозавров». стр. 263 в Lucas, S.G. and Sullivan, R.M. (ред.), Позднемеловые позвоночные из внутренних районов Запада. Бюллетень Музея естественной истории и науки Нью-Мексико 35.
- ^ а б Танке (2010); «Примечание 4», стр. 544.
- ^ Танке (2010); «Примечание 9», стр. 546.
- ^ а б c d е ж грамм час я j Хорнер, Вайсхампель и Форстер (2004); «Палеоэкология, биогеография и палеобиология», стр. 461.
- ^ а б Хорнер, Вайсхампель и Форстер (2004); «Палеоэкология, биогеография и палеобиология», стр. 463.
- ^ Хорнер, Вайсхампель и Форстер (2004); «Систематика и эволюция», страницы 457–458.
- ^ Хорнер, Вайсхампель и Форстер (2004); «Систематика и эволюция», стр. 458.
- ^ You et al. (2003); «Аннотация», стр. 347.
- ^ Кобаяши и Адзума (2003); «Аннотация», стр. 166.
- ^ Болотский и Годфройт (2004); «Аннотация», стр. 351.
- ^ Годфройт, Ли и Шан (2005); «Аннотация», стр. 697.
- ^ Prieto-Márquez et al. (2006); «Аннотация», стр. 929.
- ^ Гилпин, ДиКроче и Карпентер (2007); «Аннотация», стр. 79.
- ^ Мо и др. (2007); «Аннотация», стр. 550.
- ^ Zhao et al. (2007); «Аннотация», стр. 111.
- ^ Godefroit et al. (2008); «Аннотация», стр. 47.
- ^ Вагнер и Леман (2009); «Аннотация», стр. 605.
- ^ Переда-Субербиола и др. (2009); «Аннотация», стр. 559.
- ^ Сьюз и Аверьянов (2009); «Абстракция», стр. 2549.
- ^ Далла Веккья (2009); «Абстракция», стр. 1100.
- ^ Крусадо-Кабальеро, Переда-Субербиола и Руис-Оменьяка (2010); «Аннотация», стр. 1507.
- ^ Прието-Маркес (2010); «Аннотация», стр. 1.
- ^ Хуарес Вальери и др. (2010); «Аннотация», стр. 217.
- ^ Гейтс и др. (2011); «Аннотация», стр. 798.
- ^ Godefroit et al. (2012); «Аннотация», стр. 335.
- ^ Рамирес-Веласко и др. (2012); «Аннотация», стр. 379.
- ^ Godefroit et al. (2012); «Аннотация», стр. 438.
- ^ Кориа, Рига и Касадио (2012); «Аннотация», стр. 552.
- ^ Прието-Маркес и Браньяс (2012); «Аннотация», стр. 607.
- ^ Прието-Маркес, Чиаппе и Джоши (2012); «Аннотация», стр. 1.
- ^ Prieto-Márquez et al. (2013); "Канардия ген. ноя », стр. 5.
- ^ Белл и Бринк (2013); «Аннотация», стр. 265.
- ^ Прието-Маркес и Вагнер (2013); «Аннотация», стр. 255.
- ^ Wang et al. (2013); «Аннотация», стр. 1.
- ^ Prieto-Márquez et al. (2014); «Аннотация», стр. 1.
- ^ Гейтс и Шитц (2014); «Аннотация», стр. 798.
- ^ Xing et al. (2014); «Аннотация», стр. 1.
- ^ Гейтс и др. (2014); «Аннотация», стр. 156.
- ^ Вы, Ли и Додсон (2014); «Аннотация», стр. 73.
- ^ Шибата и Адзума (2015); «Аннотация», стр. 421.
- ^ Мори, Дракенмиллер и Эриксон (2015); "Абстрактный".
- ^ Фридман Фаулер и Хорнер (2015); временно.
- ^ Shibata et al. (2015); временно.
- ^ Xu et al. (2016); временно.
- ^ Wang et al. (2016); временно.
- ^ Prieto-Márquez et al. (2016); временно.
- ^ Терри А. Гейтс; Хишигжав Цогтбаатар; Линдси Э. Занно; Цогтбаатар Чинзориг; Махито Ватабе (2018). «Новый игуанодонт (Dinosauria: Ornithopoda) из раннего мела Монголии». PeerJ. 6: e5300. Дои:10.7717 / peerj.5300. ЧВК 6078070. PMID 30083450.
- ^ Прието-Маркес, Альберт; Фондевилла, Виктор; Sellés, Albert G .; Вагнер, Джонатан Р .; Галобарт; Ангель (2019). "Адиномозавр арканус, новый динозавр-ламбеозавр с позднемелового иберо-армориканского острова Европейского архипелага ". Меловые исследования. 96: 19–37. Дои:10.1016 / j.cretres.2018.12.002.
- ^ Цзялян Чжан; Сяолинь Ван; Цян Ван; Шуньсин Цзян; Синь Чэн; Нин Ли; Руй Цю (2019). «Новый зауролофин гадрозаврид (Dinosauria: Ornithopoda) из верхнего мела провинции Шаньдун, Китай». Anais da Academia Brasileira de Ciências. 91 (Приложение 2): e20160920. Дои:10.1590/0001-3765201720160920. PMID 28876393.
- ^ Хишигжав Цогтбаатар; Дэвид Б. Вейшампель; Дэвид С. Эванс; Махито Ватабе (2019). «Новый гадрозавроид (Dinosauria: Ornithopoda) из позднемеловой формации Байншир в пустыне Гоби (Монголия)». PLOS ONE. 14 (4): e0208480. Дои:10.1371 / journal.pone.0208480. ЧВК 6469754. PMID 30995236.
- ^ Прието-Маркес, Альберт; Вагнер, Джонатан Р .; Леман, Томас (2019). «Необычный динозавр с« лопаточным клювом »с трофическими специализациями из раннего кампана Транс-Пекос, Техас, и наследственный герб гадрозавров». Журнал систематической палеонтологии. 18 (6): 461–498. Дои:10.1080/14772019.2019.1625078. S2CID 202018197.
- ^ Виктория Ф. Кристал; Эрика С.Дж. Эванс; Генри Фрике; Ян М. Миллер; Джозеф Дж. У. Сертич (2019). «Речная гидрология позднего мела и поведение динозавров в южной части штата Юта, США: анализ стабильных изотопов биогенного карбоната». Палеогеография, палеоклиматология, палеоэкология. 516: 152–165. Дои:10.1016 / j.palaeo.2018.11.022.
- ^ Ю ‐ Гуан Чжан; Ке-Бай Ван; Шу-Цин Чен; Ди Лю; Хай Син (2019). «Остеологическая переоценка и таксономическая ревизия "Tanius laiyangensis" (Ornithischia: Hadrosauroida) из верхнего мела провинции Шаньдун, Китай ". Анатомический рекорд. 303 (4): 790–800. Дои:10.1002 / ar.24097. PMID 30773831. S2CID 73476311.
- ^ Холли Н. Вудворд (2019). "Майасаура (Dinosauria: Hadrosauridae) Остеогистология большеберцовой кости выявляет однолетние корковые сосудистые кольца у молодых годовалого возраста ». Границы науки о Земле. 7: Статья 50. Дои:10.3389 / feart.2019.00050.
- ^ Эймон Т. Дрисдейл; Франсуа Терриен; Дарла К. Зеленицкая; Дэвид Б. Вейшампель; Дэвид С. Эванс (2019). "Описание ювенильных особей Prosaurolophus maximus (Hadrosauridae: Saurolophinae) из верхнемеловой формации Bearpaw на юге Альберты, Канада, обнаруживает онтогенетические изменения в морфологии гребня ". Журнал палеонтологии позвоночных. в печати (6): e1547310. Дои:10.1080/02724634.2018.1547310. S2CID 109440173.
- ^ Пол В. Ульманн; Сурадж Х. Пандья; Рон Неллермоу (2019). «Паттерны сохранения мягких тканей и клеток в зависимости от структуры окаменелой костной ткани и глубины перекрывающих отложений на участке Стендинг Рок Хадрозавр, формация Маастрихт Хелл Крик, Южная Дакота, США». Меловые исследования. 99: 1–13. Дои:10.1016 / j.cretres.2019.02.012.
- ^ Рюдзи Такасаки; Энтони Р. Фиорилло; Ёсицугу Кобаяси; Рональд С. Тыкоски; Пол Дж. Маккарти (2019). «Первая определенная кость ламбеозаврина из костного ложа Лискомб в формации Принс-Крик в верхнемеловом периоде, Аляска, США». Научные отчеты. 9 (1): Артикульный номер 5384. Дои:10.1038 / s41598-019-41325-8. ЧВК 6440964. PMID 30926823.
- ^ Джозеф Э. Петерсон; Карсен Н. Даус (2019). "Следы кормления, приписываемые молодым тиранозавр Рекс предложить понимание онтогенетических диетических тенденций ". PeerJ. 7: e6573. Дои:10.7717 / peerj.6573. ЧВК 6404657. PMID 30863686.
Рекомендации
- Bell, P.R .; Бринк, К. С. (2013). "Kazaklambia убеждает гребень. nov., примитивный ювенильный ламбеозаврин из сантона Казахстана ». Меловые исследования. 45: 265–274. Дои:10.1016 / j.cretres.2013.05.003.
- Болотский, Ю.Л .; Годфройт, П. (2004). «Новый динозавр гадрозаврин из позднего мела Дальнего Востока России». Журнал палеонтологии позвоночных. 24 (2): 351–365. Дои:10.1671/1110. S2CID 130691286.
- Родольфо А. Кориа, Бернардо Гонсалес Рига и Сильвио Касадио (2012). "Un nuevo hadrosáurido (Dinosauria, Ornithopoda) de la Formación Allen, provincia de la Pampa, Аргентина". Амегиниана. в прессе.
- Крусадо-Кабальеро, Пенелопа; Хабьер Переда-Субербиола; Хосе Игнасио Руис-Оменьяка (2010). "Blasisaurus canudoi ген. et sp. nov., новый динозавр ламбеозавр (Hadrosauridae) из позднего мелового периода Арена (Уэска, Испания) ". Канадский журнал наук о Земле. 47 (12): 1507–1517. Дои:10.1139 / E10-081.[постоянная мертвая ссылка ]
- Далла Веккья, Ф. М. (2009). "Tethyshadros insularis, новый гадрозавроидный динозавр (Ornithischia) из верхнего мела Италии ". Журнал палеонтологии позвоночных. 29 (4): 1100–1116. Дои:10.1671/039.029.0428.
- Элизабет А. Фридман Фаулер и Джон Р. Хорнер (2015). "Новый брахилофозаврин гадрозавр (динозавр: орнитишия) с промежуточным носовым гребнем из формации Кампанской реки Джудит в северной части Монтаны". PLOS ONE. 10 (11): e0141304. Дои:10.1371 / journal.pone.0141304. ЧВК 4641681. PMID 26560175.
- Гейтс, T.A .; Хорнер, J.R .; Hanna, R.R .; Нельсон, C.R. (2011). «Новый неукрашенный гадрозаврин гадрозаврид (Dinosauria, Ornithopoda) из кампана Северной Америки». Журнал палеонтологии позвоночных. 31 (4): 798–811. Дои:10.1080/02724634.2011.577854. S2CID 8878474.
- Гейтс, Т. А .; Scheetz, R. (2014). «Новый зауролофин гадрозаврид (Dinosauria: Ornithopoda) из кампана штата Юта, Северная Америка». Журнал систематической палеонтологии. 13 (8): 711–725. Дои:10.1080/14772019.2014.950614. S2CID 129231015.
- Терри А. Гейтс, Зубайр Джинна, Кэролайн Левитт и Майкл А. Гетти (2014). «Новые образцы гадрозавридов (Dinosauria, Ornithopoda) из нижней и средней части кампанской формации Вахвип на юге Юты». У Дэвида А. Эберта; Дэвид С. Эванс (ред.). Гадрозавры: Материалы Международного симпозиума по хадрозаврам. Издательство Индианского университета. С. 156–173. ISBN 978-0-253-01385-9.CS1 maint: несколько имен: список авторов (связь)
- Гилпин, Дэвид; ДиКроче, Тони; Карпентер, Кеннет (2007). «Возможный новый базальный гадрозавр из формации кедровых гор нижнего мела Восточной Юты». В Карпентер, К. (ред.). Рога и клювы: динозавры цератопсов и орнитоподов. Блумингтон и Индианаполис: Издательство Индианского университета. С. 79–89. ISBN 978-0-253-34817-3.
- Godefroit, P .; Li, H .; Шан, С.Ю. (2005). "Новый примитивный динозавр-гадрозавроид из раннего мела Внутренней Монголии (Китайская Народная Республика)."". Comptes Rendus Palevol. 4 (8): 697–705. Дои:10.1016 / j.crpv.2005.07.004.
- Годфруа, Паскаль; Хай Шулин; Ю Тинсян; Лаутерс, Паскалин (2008). «Новые динозавры гадрозавриды из самых верхних меловых периодов северо-восточного Китая» (PDF). Acta Palaeontologica Polonica. 53 (1): 47–74. Дои:10.4202 / приложение.2008.0103. S2CID 15853629.
- Паскаль Годфруа, Франсуа Эскуйе, Юрий Л. Болотский и Паскалин Лаутерс (2012). «Новый базальный динозавр адрозавроидов из верхнего мела Казахстана». В Godefroit, P. (ed.). Динозавры Берниссара и наземные экосистемы раннего мела. Издательство Индианского университета. С. 335–358.CS1 maint: несколько имен: список авторов (связь)
- Godefroit, P .; Болотский, Ю.Л .; Лаутерс, П. (2012). Джогер, Ульрих (ред.). "Новый динозавр-зауролофин из позднего мела Дальнего Востока России". PLOS ONE. 7 (5): e36849. Дои:10.1371 / journal.pone.0036849. ЧВК 3364265. PMID 22666331.
- Хорнер, Джон Р .; Weishampel, David B .; Форстер, Кэтрин А. (2004). «Hadrosauridae». В Weishampel, D. B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. С. 438–463. ISBN 978-0520254084.
- Рубен Д. Хуарес Вальери, Хосе А. Аро, Лукас Э. Фьорелли и Хорхе О. Кальво (2010). «Новый гадрозавроид (Dinosauria: Ornithopoda) из формации Аллен (поздний мел) в Патагонии, Аргентина» (PDF). Revista del Museo Argentino de Ciencias Naturales N.s. 11 (2): 217–231. Архивировано из оригинал (PDF) на 2011-09-03.CS1 maint: несколько имен: список авторов (связь)
- Кобаяши, Ю .; Адзума, Ю. (2003). «Новый игуанодонтиан (Dinosauria; Ornithopoda), из нижнемеловой формации Китадани префектуры Фукуи, Япония». Журнал палеонтологии позвоночных. 23 (1): 166–175. Дои:10.1671 / 0272-4634 (2003) 23 [166: anidof] 2.0.co; 2.
- Лукас, Спенсер Г. (2001-11-15). Китайские ископаемые позвоночные. Издательство Колумбийского университета. п. 320 (стр. 34). ISBN 978-0231084826.
- Mo J .; Zhao Z .; Wang W .; Сюй X. (2007). «Первый динозавр гадрозаврид из южного Китая». Acta Geologica Sinica (английское издание). 81 (4): 550–554. Дои:10.1111 / j.1755-6724.2007.tb00978.x.
- Мори, Хироцугу; Дракенмиллер, Патрик С. и Эриксон, Грегори М. (2015). «Новый арктический гадрозаврид из формации Принс-Крик (нижний маастрихт) на севере Аляски». Acta Palaeontologica Polonica. 61 (В прессе). Дои:10.4202 / app.00152.2015.
- Переда-Субербиола, Ксабье; Хосе Игнасио Канудо; Пенелопа Крусадо-Кабальеро; Хосе Луис Барко; Ньевес Лопес-Мартинес; Ориол Омс; Хосе Игнасио Руис-Оменьяка (2009). «Последние динозавры-гадрозавриды в Европе: новый ламбеозаврин из самых верхних меловых периодов Арена (Уэска, Испания)» (PDF). Comptes Rendus Palevol. 8 (6): 559–572. Дои:10.1016 / j.crpv.2009.05.002.
- Prieto-Marquez, A .; Gaete, R .; Rivas, G .; Galobart, Á .; Боада, М. (2006). "Гадрозавроидные динозавры из позднего мела Испании: Pararhabdodon isonensis пересмотренный и Кутализавр kohlerorum, ген. et sp. ноя ". Журнал палеонтологии позвоночных. 26 (4): 929–943. Дои:10.1671 / 0272-4634 (2006) 26 [929: hdftlc] 2.0.co; 2.
- Прието-Маркес, Альберт (2010). "Glishades ericksoni, новый гадрозавроид (Dinosauria: Ornithopoda) из позднего мела Северной Америки » (PDF). Zootaxa. 2452: 1–17. Дои:10.11646 / zootaxa.2452.1.1.
- Прието-Маркес, Альберт; Серрано Браньяс, Клаудиа Инес (2012). "Latirhinus uitstlani, «широконосый» зауролофин гадрозаврид (Dinosauria, Ornithopoda) из позднего кампана (меловой период) северной Мексики ". Историческая биология. 24 (6): 607–619. Дои:10.1080/08912963.2012.671311. S2CID 128964878.
- Prieto-Márquez, A .; Chiappe, L.M .; Джоши, С. Х. (2012). Додсон, Питер (ред.). "Динозавр-ламбеозавр Magnapaulia laticaudus из позднего мела Нижней Калифорнии, Северо-Западная Мексика ". PLOS ONE. 7 (6): e38207. Дои:10.1371 / journal.pone.0038207. ЧВК 3373519. PMID 22719869.
- Prieto-Márquez, A .; Dalla Vecchia, F.M .; Gaete, R .; Галобарт, А. (2013). Додсон, Питер (ред.). «Разнообразие, родство и биогеография динозавров ламбеозавров с Европейского архипелага с описанием нового аралозаврина. Canardia garonnensis". PLOS ONE. 8 (7): e69835. Дои:10.1371 / journal.pone.0069835. ЧВК 3724916. PMID 23922815.
- Прието-Маркес, Альберт; Вагнер, Джонатан Р .; Белл, Фил Р .; Чиаппе, Луис М. (2014). "Поздно выживший динозавр с утиным клювом" Августинолоф из верхнего маастрихта западной части Северной Америки и эволюция гребня в Saurolophini ». Геологический журнал. 152 (2): 225–241. Дои:10.1017 / S0016756814000284.
- Прието-Маркес, Альберт; Эриксон, Грегори М .; Эберсол, июн А. (2016). «Примитивный гадрозаврид из юго-востока Северной Америки и происхождение и ранняя эволюция динозавров с утиным клювом». Журнал палеонтологии позвоночных. 36 (2): e1054495. Дои:10.1080/02724634.2015.1054495. S2CID 86032549.
- Прието-Маркес, А.; Вагнер, Дж. (2013). «Новый вид зауролофиновых динозавров гадрозаврид из позднего мела Тихоокеанского побережья Северной Америки». Acta Palaeontologica Polonica. 58 (2): 255–268. Дои:10.4202 / app.2011.0049.
- Анхель Алехандро Рамирес-Веласко, Мулуд Бенамми, Альберт Прието-Маркес, Хесус Альварадо Ортега и Рене Эрнандес-Ривера (2012). "Huehuecanauhtlus tiquichensis, новый гадрозавроидный динозавр (Ornithischia: Ornithopoda) из сантона (поздний мел) штата Мичоакан, Мексика ". Канадский журнал наук о Земле. 49 (2): 379–395. Дои:10.1139 / e11-062.CS1 maint: несколько имен: список авторов (связь)
- Масатеру Сибата, Пратуенг Джинтасакул, Йоичи Адзума и Хай-Лу Ю (2015). «Новый базальный динозавр-адрозавроид из формации Кок-Круат нижнего мела в провинции Накхонратчасима, Северо-Восточный Таиланд». PLOS ONE. 10 (12): e0145904. Дои:10.1371 / journal.pone.0145904. ЧВК 4696827. PMID 26716981.CS1 maint: несколько имен: список авторов (связь)
- Шибата, Масатеру; Адзума, Йоичи (2015). «Новый базальный гадрозавроид (Dinosauria: Ornithopoda) из формации Китадани нижнего мела, Фукуи, центральная Япония» (PDF). Zootaxa. 3914 (4): 421–40. Дои:10.11646 / zootaxa.3914.4.3. PMID 25661952.
- Сьюз, Ганс-Дитер; Аверьянов, Александр (2009). «Новый базальный гадрозавроидный динозавр из позднего мела Узбекистана и раннее излучение утконосых динозавров». Труды Королевского общества B: биологические науки. 276 (1667): 2549–2555. Дои:10.1098 / rspb.2009.0229. ЧВК 2686654. PMID 19386651.
- Танке, Д. Х. (2010). «Потерянный на виду: повторное открытие пропавших без вести Эоцератопсов Уильяма Э. Катлера». В Ryan, M. J .; Chinnery-Allgeier, B.J .; Эберт, Д. А. (ред.). Новые взгляды на рогатых динозавров: Цератопсовский симпозиум Королевского музея Тиррелла. Жизнь прошлого. Блумингтон: Издательство Индианского университета. С. 541–550. ISBN 978-0253353580.
- Вагнер, Джонатан Р .; Леман, Томас М. (2009). «Загадочный новый ламбеозавр гадрозавр (рептилии: динозавры) из верхней сланцевой пачки кампанской формации Агуджа в Транс-Пекос, штат Техас». Журнал палеонтологии позвоночных. 29 (2): 605–611. Дои:10.1671/039.029.0208. S2CID 128555861.
- Weishampel, David B .; Янг, Л. (1996). Динозавры восточного побережья. Издательство Университета Джона Хопкинса.
- Wang, R. F .; Вы, Х.Л .; Xu, S.C .; Wang, S. Z .; Yi, J .; Xie, L.J .; Jia, L .; Ли, Ю. X. (2013). Эванс, Дэвид С. (ред.). «Новый динозавр-адрозавроид из раннего и позднего мелового периода провинции Шаньси, Китай». PLOS ONE. 8 (10): e77058. Дои:10.1371 / journal.pone.0077058. ЧВК 3800054. PMID 24204734.
- Жунь-Фу Ван, Хай-Лу Ю, Суо-Чжу Ван, Ши-Чао Сюй, Цзянь И, Ли-Хуан Се, Лэй Цзя и Хай Син (2016). «Второй гадрозавроидный динозавр из раннего позднего мела Цзоюнь, провинция Шаньси, Китай». Историческая биология: международный журнал палеобиологии. 29: 1–8. Дои:10.1080/08912963.2015.1118688. S2CID 130536621.CS1 maint: несколько имен: список авторов (связь)
- Xing, H .; Wang, D .; Han, F .; Салливан, К .; Ma, Q .; Привет.; Hone, D. W. E .; Ян, Р .; Du, F .; Сюй, X. (2014). «Новый базальный динозавр-адрозавроид (Dinosauria: Ornithopoda) с переходными особенностями из позднего мелового периода в провинции Хэнань, Китай». PLOS ONE. 9 (6): e98821. Дои:10.1371 / journal.pone.0098821. ЧВК 4047018. PMID 24901454.
- Xu, S-C .; Вы, H-L .; Wang, JW .; Ван, С-З .; Yi, J .; Цзя, Л. (2016). «Новый гадрозавроидный динозавр из позднего мела Тяньчжэня, провинция Шаньси, Китай». Позвоночные PalAsiatica. 54 (1): 67–78.
- Вы, Х.-И .; Li, D.-Q .; Додсон, П. (2014). "Gongpoquansaurus mazongshanensis (Лю, 1997) гребешок. ноя (Ornithischia: Hadrosauroida) из раннего мела провинции Ганьсу, Северо-Западный Китай ». In Eberth, David A .; Evans, David C. (ред.). Гадрозавры. Издательство Индианского университета. С. 73–76. ISBN 978-0-253-01390-3.
- Ты, Ло; Шубин, Витмер; Тан; Тан (2003). «Самый ранний известный динозавр с утконосом из отложений позднего раннего мелового периода на северо-западе Китая и эволюция гадрозаврид». Меловые исследования. 24 (3): 347–353. Дои:10.1016 / s0195-6671 (03) 00048-х.
- Чжао, X .; Li, D .; Вешать.; Hao, H .; Лю, Ф .; Li, L .; Фанг, X. (2007). "Zhuchengosaurus maximus из провинции Шаньдун ». Acta Geoscientia Sinica. 28 (2): 111–122. Дои:10.1007 / s10114-005-0808-х. S2CID 119700784.
внешняя ссылка
 СМИ, связанные с Hadrosauroida в Wikimedia Commons
СМИ, связанные с Hadrosauroida в Wikimedia Commons